There
are
a number
of
situations
in developing
countries
in which pasture
plays
a significant
role
in the
animal production
system
as follows:
Traditional grazing systems that include:
grazing
on non-arable
land
communal
grazing
close
to
villages
communal
and traditional
grazing
of nomadic
routes
and pastures
Large and
extensive ranches
In
this
chapter,
the
application
of
basic
principles
of feeding
ruminants
is
discussed
in relation
to the
different
pasture-based
systems
that
have
evolved.
9.1.1 Extensive ranching
While
large
ranches
account
for considerable
areas
in certain
countries,
especially
in Latin
America
and
in parts of
Africa and Indonesia,
the
number
of
people
that
benefit
from
these activities
is
relatively
small.
A typical
example
is
Brazil,
where
large
areas
of
the Amazonian
region
have been
converted
from
forest
into grazing
land;
individual
farms often
reach
one million
hectares.
The economic
advantages
of introducing
technological
innovations
are extremely
limited
as it
is
usually
more
profitable
to
farm large
areas
extensively
than
to intensify
production
within
the area.
The
most
important
issue
is that in most developing
countries
this kind of exploitation
involves
a relatively
small proportion
of both the animals and people
in the country. In Asia and
Africa
70 to 90%
of the
ruminant livestock
is kept
by small
farmers
who
own
one
to five
animals.
This equates
to some
120 million cattle
and 180
million
sheep
and goats
in Africa
and 400 million
cattle and 500 million
sheep
and
goats in Asia. There
are
twice
as many ruminant livestock
in Africa or
Asia as in Europe.
The
owners
of
these livestock
represent
the major target
for development
aid and yet
these
people
have probably benefited
least
from technical
innovations.
Pastures
in the
tropics
and
subtropics
grow rapidly during periods
of heavy
rainfall
and high temperatures,
leading
to
mature
pasture
plants containing
high levels
of
cell
wall constituents.
Pastures
are young
and
green
(ie. high
in
N, soluble
carbohydrates
and
fats and
of high
digestibility)
for
only
short
periods.
The
nutritive
value
of the
pastures decreases
rapidly
with maturity
and,
during
the dry
seasons,
the available
feed
is of
low digestibility
and low
in total
N. The
major
factor
that limits animal production
from
these
pastures
is the
fact that the
animals lose
weight
in the
dry
season due
to
the
nutritional imbalance
in the
feed
available
(see Figure
9.1).
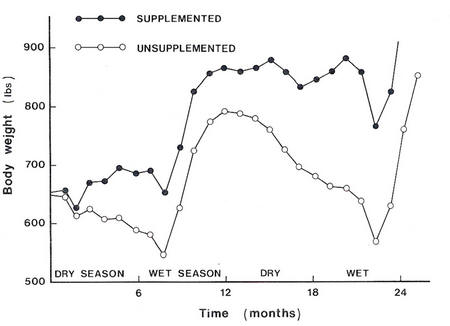
Figure
9.1 : Monthly live
weight
change
of cattle
given
no supplements
or supplemented
with
groundnut cake
(Source:
Murray
et al.
1936)
In most
developing
countries pasture
management
is restricted
largely
to grazing
or burning or
a combination
of the two.
The pastures
usually· consist of indigenous
species
which, in general,
are
low
in protein,
have
relative
low
digestibility
and, depending
on
soil
type,
low
concentrations
of
minerals.
Stobbs (1969)
suggested
that the lower
animal productivity
from
tropical
pastures
than from
temperate
pastures
may be
related to the more
erect
growth habits of
most tropical
grasses
and legumes.
The
low proportion
ofleafy
material
in these
pastures
appears
to limit harvest
ability and intake
of the
plants by grazing animals.
As the
grasses flower
and mature,
forage
quality declines
because
of
translocation
of
soluble carbohydrates
from
the stem
and leaves
to the inflorescence,
thereby
increasing the relative
proportion
oflignified
cell
walls
in the leaves
and stems. The
digesti bili
ty of tropical pasture
species
declines
much more rapidly
than that
of temperate
species. Flowering
in legumes
is not usually
associated
with such large
changes in
nutritive
value
as in grasses despite
leaf
loss
due to senescence.
In
the
heavily
populated Third World
countries,
the land
available for grazing
is limited and is frequently
of low fertility.
There is considerable
scope for
increasing
animal productivity from grass land through
management.
The options include:
Manipulation of stocking rate
Burning
Introduction of legumes and grasses by oversowing
Fertilizer application
Pasture renovation and reseeding with introduced grasses
and legumes
Establishment of protein banks (eg. small areas of legumes)
Strategic supplementation
A major problem
is how to apply
management
strategies on communally
owned
land. For
instance,
grazing pressure
is rarely
controlled
and
fertiliser
is almost
never
applied.
Agronomic
aspects of pasture improvement are
outside
the scope
of this
book. The
discussion
is restricted
to strategies
that
are specifically
aimed
at improving
the balance of nutrients
for animal productivity,
ego strategic
supplementation,
the introduction
of legumes
and use
of fertiliser
to increase
the availability
of critical
nutrients
in the
diets of grazing
or tethered ruminants
on pasture-based
diets.
9.1.2
Improving
pastures
with legumes
The role
of legumes in improving
pasture quality and animal
production from both temperate
(Ulyatt
1980) and tropical pastures (Milford
1967) is indisputable.
Animal
production is nearly always greater from legume-based
pastures than from
pure grass pastures
(Mannetje
1984; Thomson
1977; Walker
1987). A most important
attribute
oflegumes is that
their
digestibility
declines more slowly
with maturity
and environmental
temperature than does that of
grasses
(Minson 1980).
The use of adapted
legumes
to improve the
quantity
and quality
of forage from tropical pastures has been
demonstrated
in many
environments. In
some, the introduction of legumes
to native pastures
has substantially
increased
animal
production without
any fertiliser
input. For instance,
Clatworthy
and Holland (1979) observed a 53% increase
in liveweight
gain of cattle
on
Hyparrhenia
pastures
in
Zimbabwe when
legumes
were introduced.
Stobbs
(1966, 1969) reported
increases
of 11 to 49%
from
similar pastures in
Uganda.
Modest inputs of fertiliser
in addition
to introducing legumes
have further
increased
liveweight
gain
(see Shaw
and
Mannetje
1970)'
and, more importantly,
increased
the reproductive
rate of cattle (see Holroyd
et al.
1977).
The increase in animal production
is not only due
to higher bodyweight
gains but is also
due to higher carrying capacity,
which
may increase
two- to six-fold.
Beef production
of
between
260kg
and 500kg liveweight
per hectare
per year has
been
obtained
from
sown
tropical
grass/legume
pastures
(Mannetje
1984). Milk
production
from tropical
pastures and the composition
of
the
milk
produced
are
certainly
influenced
by introducing
legumes.
The increase
in
animal
production
from
including
legumes
in tropical
pastures
is
not
attributable
to a single
factor.
There
are complex
interactions
between
the extra
dry
matter available
for
grazing
and
the
increased
N content
of the herbage
selected.
There
are
several
other
reasons
why
legumes
may increase
animal
productivity.
Legumes
increase
the dry matter
production
of
the associated
grasses due
to higher
soil
N. Legumes
have
higher
mineral
concentrations
than grasses
and there
is the
potential
contribution of
seed
retained
on
the plant.
The
seeds
of
many
legume
species
contain
up to
45%
protein,
up to
0.8%
phosphorus,
0.3 to
0.5%
sulphur
and have a
digestibility
of 50-80%.
They
have
also
a high lipid
content
which may
be
of considerable
importance
where
the
grasses
are
mature
and
dry.
Legumes
also
tend to
remain
green
into the
dry season,
thus prolonging
the
grazing period.
9.1.3
Management
of legumes
In pastures
Introducing
legumes
into
pastures
is
one
option
for
increasing
animal
productivity
(see
Table
9.1).
Table
9.1:
Lambs grow faster on legume
than on
grass pasture
Grass
Mixed grass/legume
Legume
USA
127
172
213
New Zealand
57
172
New
Zealand
122
254
New Zealand
37
128
191
Australia
82
159
Australia
83
148
Source:
Archer
(1980).
However,
the
method
of
introduction,
the
species
introduced
and the
management
needed
to retain
the legumes
in the
pasture differ
according
to
soil
and climate.
For these
reasons
the
transfer
of
technology
has
often
failed
even
within a country
(Mannetje
1982).
Management
is a major
constraint in the
humid tropics.
For
instance,
in research
reported
from
Indonesia
(Blair et
al.
1985)
marked
changes
in legume
content
in sown
pastures
were
observed
with different
management
systems.
Grazing progressively
reduced
the proportion
of legumes
in the
pasture from 35 to
5%, with associated
increases
in the
invasion
by weeds
and by Imperata
cyclindrica,
which is particularly
difficult
to manage.
By
contrast,
grazing did not
affect
the
proportion
of
native
legume
in the
indigenous
pastures,
but the
proportion
of
legumes
in these
pastures
was initially low.
This
bears
out Mannetje's
viewpoint
that:
"Sown grass/
legume
pastures
are
persistent
and productive
provided
they
consist
of
adapted
species,
are
adequately
fertilized
and are
not
overgrazed"
(Mannetje
1984)
All these
factors are
almost
impossible
to ensure
in tropical
developing
countries.
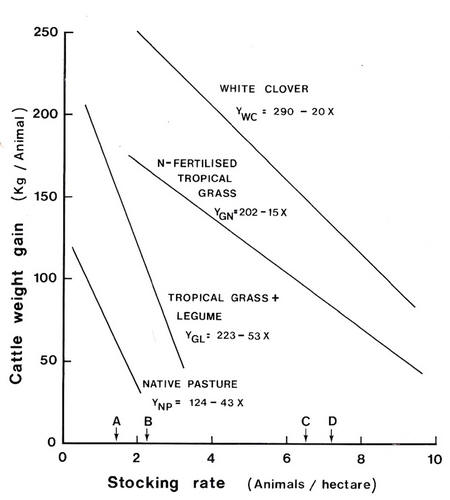
The
data in Table
9.2 and Figure
9.2
show
animal
production
from
pastures
in three
contrasting
climatic
environments.
Figure
9.2: Cattle
growth
on
pasture
is a
function
of
pasture
type,
fertiliser
application
and legume
content.
Productivity
per
unit area
is maximised
for the
different
pastures
at different
stocking
rates: 89kg/ha
for
native
pasture,
223kg/ha
for
tropical grass with
legume,
682
kg/ha
for tropical
grass
with fertiliser
and on temperate
pasture
(clover)
1051
kg/ha
(Source:
Walker
1987).
Table
9.2:
Cattle
production
from
native
(NP)
and
imp1>oved
pasture
(Imp)
with and without
legume
and
N fertiliser.
The
genetic
potential
for
growth
of
the
animals
was presumed
to be 0.6,
1.2
and
1.2kg/d
for
the
three
Rainfall
areas
A, Band
C respectively
Grass only
Improved pasture
NP
Imp
+ legume
+N
Rainfall
(A)
2Scm
Stocking
rate, /ha
#
4
8
LW
gain, g/d
160
280
360
LW
gain/ha
(kg/yr)
410
1050
Rainfall
(B)
32cm
Stocking
rate, /ha
4.2
4.2
4.2
LW
gain, g/d
240
300
380
LW
gain/ha
(kg/yr)
367
452
595
Rainfall
(C)
7.2cm
Stocking
rate, /ha
0.3
0.7
1.1
1.0
LW
gain, g/d
310
280
420
420
LW
gain/ha
(kg/yr)
30
76
167
167
Source:
Mannetje
(1982).
.#Village:
conditions
The
data indicate
that:
Although
average daily gain of animals in all rainfall areas can be increased by
introducing legumes and/or fertilizing the pastures, animal productivity
is still
well
below
the genetic
potential
Animal
production
per
hectare
can be
increased
substantially
by
increasing
stocking
rate
according
to the
increased
availability
of forage
dry
matter
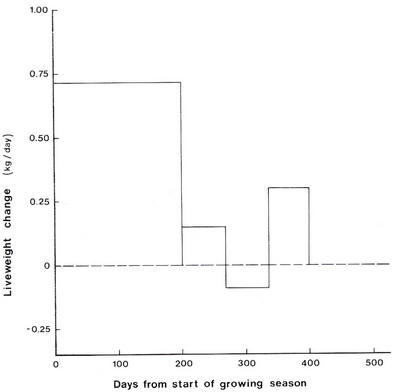
The
low production
in Australia
is
largely
attributable
to
seasonal
differences
in rainfall;
animals
grow
about
three
times
faster
on
green
and
fertilized
pasture
than
on
the
dry
pasture
during
the
rest
of the
year
(see
Figure
9.3).
Figure
9.3:
Seasonal
variation
in
liveweight
gain response
of
cattlegrazing
Cenchrus
ciliaris(Source:
Mannetje
1982).
Apart from
the
agronomic
problems
of retaining
legumes,
it is
apparent
that
their
presence
in
the
pasture
does
not overcome
the
inherent
nutritional
constraints.
The
relatively
small increases
in productivity
when
legumes
are
included
in a pasture,
compared
with
the
responses
that
can
be
achieved
on
dry
pastures
when
the
animals
are
supplemented
with
meals
providing
bypass
protein
(and
minerals,
particularly
phosphorus)
(see
Table
9.7),
indicate
that
the
legumes
are
not
providing
the
critical
bypass
nutrients
but
merely
fermentable
N.
9.1.4
Use
of fertilisers to Improve grassland
On soils
of low
fertility,
fertilisers
undoubtedly
increase
dry
matter
production
per
hectare.
The
most
widely
used
fertilisers
are
superphosphate
(S
and P)
and nitrogen.
The
increase
in animal
productivity
from
pastures
fertilized
with
superphosphate
is especially
noticeable
in the
tropics,
where
phosphorusdeficient
soils
are
common.
In general,
phosphorus
fertilisers
are
used
at low
rates
to promote
legume
growth
and
thus
increase
pasture
productivity.
The
use
of nitrogen
fertilisers
to
maximise
herbage
production
has generally
been
applied
in intensively
managed
systems
in which
high
value
products
(eg.
milk)
can justify
the
economic
cost.
Table
9.3:
Indicative
amounts
of feed
biomass
required
to fatten
cattle
to
market
weight
(ie,
100-4
OOkg
liveweight)
at pasture
assuming
a
range
of growth
rates
and approximate
conversion
ratios
(FeR
in
kg DM/kg
L Wt
gain).
Feed
biomass
LWt
gain
(kg/d)
FeR (kg/kg)
(tonnes
DM)*
.2
25
7.5
.4
15 4.5
. 6
12
3.6
.8
9 2.7
1.0
6
1.8
9.1.5
Pasture improvement and stocking rate
In
developed
countries
with
mainly
pastoral
economies
(eg.
New
Zealand
and Australia),
the
approach
to improving
animal
productivity
has
been
mainly
to maximise
production
per
unit
area,
which
has
usually
meant
sacrificing
productivity
per
animal
(Figure
9.2).
The
next
developments
must
aim at increasing
individual
animal
productivity
by
identifying
the
major
imbalances
in nutrients
absorbed
by grazing
ruminants
and
using
supplements
in
order
to
correct
the
deficiencies.
The
data
in Table
9.3,
derived
from
assumed
rates
of
growth
and
feed
conversion,
indicate
the
differences
in the
quantities
of feed
biomass
needed
to raise
animals
from
weaning
to
market
weight
(ie.
from
100 to
400kg)
at different
growth
rates,
and
emphasise
the
inefficiencies
of
a strategy
aimed
at increasing
productivity
per
unit area
at the
expense
of
individual
animal
productivity.
Table
9.3:
Indicative
amounts
of feed
biomass
required
to fatten
cattle
to
market
weight
(ie,
100-4
OOkg
liveweight)
at pasture
assuming
a range
of growth
rates
and approximate
conversion
ratios
(FeR
in
kg DM/kg
L Wt
gain
LWt
gain
(kg/d)
FCR
(kg/kg)
Feed
biomass
(tonnes
DM)#
0.2
25
7.5
0.4
15
4.5
0.6
12
3.6
0.8
9
2.7
1.0
6
1.8
# From 100
to 400 kg LW
The
foregoing
discussion
has highlighted
the
issues
that
should
be addressed
in seeking
to
increase
animal
productivity
on pasture,
including
the
capacity
of
the
forage
to
support
efficient
rumen
function
and to
provide
additional
bypass
nutrients
and
long chain
fatty
acids.
These
critical
factors
are
not
defined
by chemical
analysis
alone.
9.2 Nutritive value
9.2.1
Supplementation of ruminants on green pastures
The
major
nutritional
differences
between
tropical
and
temperate
pastures
are
that
temperate
pastures
are
usually
higher
in protein
and soluble
sugars
and lower
in cell
wall
components
and
therefore
are
more
digestible.
Even
on
temperate
pastures
that
are
of
high
digestibility
and contain
legumes,
growth
rates of
lambs
are
usually
considerably
lower
than
the
genetic
potential
and sometimes
less
than
that obtained
on
correctly
supplemented
crop
residues
(see
Chapter
8).
Growth
rates
and milk production
of
cattle and
sheep
on
tropical
pastures
are
usually
lower
than those of
animals
grazing temperate
green
pastures
(Walker
1987).
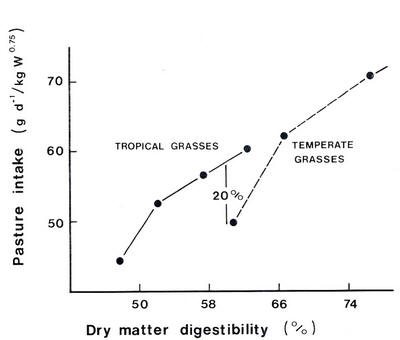
Figure
9.4
illustrates
the
relationship
between
voluntary
feed
intake
and
digestibility
for
a wide
range
of
tropical
and
temperate
pastures.
At a digestibility
of about
60%,
which
is
high
for
tropical
pastures,
the
forage
intake
of sheep
is
greater
on
tropical
pasture
than
on
temperate
pasture
of
the
same
digestibility
(Minson
1980).
Figure
9.4:
Relationship
between
voluntary
intake
and dl'y
matter
digestibllity
fol'
a range
of
tropical
and temperate
grasses
(SoU7'ce:
Minson
1980,
1982)
In
the
humid
tropics,
in order
to maintain
the digestibility
of
pasture
at a relatively
high
level,
grazing
pressure
should
be
increased
to
a level
at which
the pastures
are
kept
young
and vegetative.
However,
this
reduces
the
amount
of feed
on
offer,
which
reduces
intake.
The
greatest
risk
is that
the animal
will not have
enough
feed
for
maintenance
if pasture
growth rate
slows
because
of drought.
In these
conditions,
appetite
may
be
limited
also
by
fatigue
resulting
from the
work
needed
to gather
the pasture
available
(McClymont
1967).
On the other
hand,
in many
tropical
livestock
systems,
animals
are
corralled
at night
and are
fed
cereal straws
and
other
residues
that
are
low
in fermentable
N and
protein.
Consuming
small amounts
of leafy oasture
during
the
day
would
provide
the
nutrients needed
to support
the rumen
ecosystem,
and hence
permit more
efficient
use of
the crop
residues
fed at night.
If stocking
intensity
is
low
on
tropical
pastures,
the proportion
of
mature
vegetation
in the
pasture
increases rapidly.
The
mature
vegetation
is often
unpalatable and,
after
a certain
stage of
maturity,
the
the pasture is either
not grazed
or
only
the
new
shoots
are grazed. Under
these
circumstances
the
cattle
graze
a much smaller
area of
land than indicated
by stocking
density and
this
area
is heavily
over
grazed,
resulting
in low feed intake
and low
productivity.
The
custom
of burning grasslands
presumably
arose
from
this tendency
of ruminants
to
concentrate
their
grazing
on small
areas
of land. Burning
the ungrazed
areas
induces regrowth
and encourages
grazing
over
a widerare
and also
controls
tick
infestation
to some
extent
(Chapter
10).
At high
stocking
rates
on
green
pastures,
production
is probabably
limited
by availability
of herbage
and therefore a
supplementary
feeding
programme
should
aim to supply
a balanced
feed.
Where
pasture
availability is
not
limiting,
productivity
is
probably
constrained by low
feed
intake
because
of low
digestibility
and/or
imbalance
of absorbed
nutrients
in relation
to the animals'
requirements.
Supplementary
feeding
therefore
should aim
to balance
the
absorbed
nutrients to stimulate
or maintain,
rather
than depress,the
intake of the basal
(low
cost)
pasture.
The
development
of a
rational
supplementary
feeding
practice
requires a knowledge
of the
factors
that
influence
the
balance of nutrients
from
rumen
fermentation
according
to the principles
discussed
in Chapters
3, 4,
5 and
7.
9.3 Feed intake at pasture
Apptetite control
in grazing
ruminants
is
complex
but the most important
factors
are:
The balance
of protein
relative
to
energy in the
products available
for
absorption
The rate of digestion of feed
The
rate
of comminution
of
feed
particles
to
sizes
that can
readily
flow
from
the rumen.
Voluntary
intake
of tropical
pastures
by
ruminants
varies
from
30 to
85g/kg0.75
per day
(1 to 3%
of body
weight)
according
to the
pasture
and time
of the
season.
Intake
on tropical
pastures
is generally
low
compared
with that
recorded
for
immature
temperate
pastures.
For
example,
Minson
(1980)
reported
an
intake
of 140g/kg
0.75per
day
for
a temperate
pasture.
This
suggests
that
there
are
basic
differences
between
tropical
and temperate
pastures.
This
may be
in
the
availability
of nitrogen
(both
fermentable
and
bypass
protein
N) and minerals
(depending
on soil
type),
which
are
generally
higher
in temperate
than
tropical
plants.
The
primary
factors
limiting
feed
intake
on pasture
will determine
the
supplements
to
be
used.
If digestibility
of
the pasture
is
low
and distension
of
the rumen
is limiting feed
intake, the
supplement
must have a
'low
rumen
load'
(ie.
either
rapidly
fermented
or
is rapidly
passed
to
the lower
tract,
where
it is
readily
digested).
Soluble
sugars
(eg.
molasses),
high
digestibility
seed
grains such
as lupins
and whole
cottonseed,
and high
digestibility
byproducts
such
as citrus pulp and sugar
beet
pulp might
be used
to increase
energy
intake
and provide
extra digestible
protein
and VFA energy
through
fermentation
and growth
of
micro-organisms
in the
rumen.
Feeding
molasses
to
cattle
on green
pastures
can
be highly
beneficial
and,
provided
that the
pasture
has
a high
N content,
additional
fermentable
N is
not
needed.
But increasing
the content
of
soluble
sugars,
or
other
rapidly
fermented
carbohydrates
such as grain,
in a forage
diet
may encourage
the
growth
of protozoa
in the
rumen,
which
may reduce
the
amount
of microbial
protein
available
to
the
animal
and
may also
decrease
fungal
colonisation
of plant
cell
walls
and thus
decrease
the
rate
of digestion
(see Chapters
3 and 5).
This
kind of
supplementation
can
also
depress
cellulolysis
by
reducing
the
rumen
pH or
because
of a "soluble
carbohydrate"
effect
(Mould et
al1983/84).
Chopping
et
al. (1970),
working
with dairy cows
grazing irrigated
and
fertilized
pangola grass,
showed
that feeding
3-4 kg
of molasses
daily to
cows
increased
milk production
by up to 2.5 litres
per
day.
At higher
levels
of molasses
feeding
there
was a marked
decline
in response,
which did not
occur
when
the animals were
given
cereal
grain as the supplement.
However,
generally
poorer
results
for such
molasses
supplementation
were
reported
by Preston
and Willis
(1974)
for
cattle
in Cuba.
The
difference
between
the two
studies
was
probably
due
to
the
higher
protein
content
of the
irrigated
pasture.
9.4 Supplementation
9.4.1 General
supplementation practices
Although
productivity
per
animal
from
tropical
pasture
is usually
less
than that from
temperate
pastures,
it is
almost
always
uneconomic
to
feed
supplements.
However,
as the pasture
matures
and turns into
dry
standing forage,
individual animal productivity
declines
and bodyweight losses
are
often
excessive
(see
Figure
9.1).
The digestibility
of
dry standing
forage
deteriorates
with time and
with the
incidence
ofdews
or light rain
(which
promote
saprophyte
growth
on
the
pasture).
Often
these
standing
hays are
less
than
45%
digestible
and
exceedingly
low
in
soluble
carbohydrates
and total
protein
(usually
around
0.5%
N).
They
are
not
markedly
different
in composition
from
many
cereal
straws.
Depending
on
the
soil
and
the
incidence
of rain,
they
are
often
low
in particular
mineral
elements
and may
contain
oxalates
which
form
highly
insoluble
calcium
salts
in
the
rumen.
The
constraints
to
ruminant
production
on
such
feeds
are
obvious
and a clear
set
of
principles
for
supplementary
feeding
can be
suggested.
In
considering
supplementary
feeding
practices,
the
first step
must
be
to
establish
the order
of likely
constraints.
Listed
below
are the
factors
that
are
believed
to
be
the
main limitations
to
the
utilisation
of dry
feed
by
ruminants,
which
are
similar to
those
discussed
for
straw-based
diets.
At
times
any
one
of
these
may
be
the
first
limiting factor
to
production:
Insufficient fermentable Nand S in the diet to support efficient
rumen function
Low rate of flow of digesta out of the rumen, creating distension
and limiting feed intake
An imbalance in the protein-to-energy ratio in absorbed products,
depending on the physiological state of the animal and parasitic
infection
Lack of overall availability of protein and energy because of low
feed intake
Mineral deficiencies affecting the efficiency of growth of rumen
organisms and the animal.
Many
of
these
factors
are
interrelated.
For
instance
a
lower-than-optimum
fermentable
N content
in the
diet
may
decrease
digestibility
as
well
as
resulting
in
a low
ratio
of
amino
acids
to
energy
in
the
absorbed
nutrients.
Increasing
the
availability
of fermentable
N
increases
the
digestibility
and the
protein-to-energy
ratio
in
absorbed
products
because
of
increased
efficiency
of fermentation
in the
rumen,
and both
effects
lead
to
increased
intake
of pasture.
9.4.2 Mineral supplements
Mineral
deficiencies
are
not
easily
predicted
because
of the
variability
in mineral
content
of
pastures,
which depends
on
soil,
climate, previous
stocking
history and fertiliser
application. Minerals
can usually
be added
to a
diet
relatively
cheaply,
and in practice
minerals
can be
provided
by
a supplement
or block lick.
It is
emphasised
that mineral
requirements
are
low
in animals
that
are
only
maintaining
weight,
and mineral
deficiencies
will
only
become
apparent
when other
limiting
factors
have
been
removed
and the
animal
has
the
potential
to
grow
or
produce.
Low
cost improvements
of native
pastures
has
centred
in Australia
on
augmentation
with legumes
and application
of
phosphate
fertiliser.
However,
it has been
shown
that
where
the
legumes
can
be
sustained
without
fertiliser
application
then
it is
more
economic
to supplement
the
animals
with
phosphorus
than to apply
it
to
the
soil
(see
Walker
1987).
There
is
a vast
amount
of
literature
on mineral
deficiencies
in
grazing
livestock
(see
Butterworth
1985).
Often,
recommendations
for mineral supplementation
are
made
without
considering
other
more
limiting constraints.
Thus,
if animals
are
short
of feed,
or there
is
an imbalance
of
essential
nutrients
(eg.
ammonia
for
rumen
fermentation),
correcting
a mineral
imbalance
is
unlikely
to
affect
animal performance
(see Figure
8.26).
The
high
levels
of
oxalates
in tropical
pastures
may
reduce
the
availability
of
calcium
for
absorption,
which at
times
could limit production.
Horses
on
tropical
pastures,
unless
supplemented,
exhibit
symptoms
of
calcium
deficiency,
including
the
condition
'big
head'
(McKenzie
et
al 1981),
but ruminants
that are
adapted
to such
diets
have
bacteria
in the
rumen
that
can
degrade
oxalate.
Blaney
et
al (1982)
demonstrated
that
the
availability
of
calcium
from tropical
grasses,
in
which
oxalate
is
present,
is
about
60%
of that
of
grasses
containing
little
oxalate.
There
are
reports
of
responses
in
cattle
on dry
subtropical
pastures
to sodium
chloride
supplements
(Murphy
and
Plasto1973;
see
also
Butterworth
1985). Under
these
circumstances
of apparent
sodium
deficiency,
it
is
claimed
that
supplementation
with
urea
is
ineffective
unless
accompanied
by sodium
supplementation.
Often
supplements
such as
poultry
litter
or bone
meal
can provide
the
major
minerals
required
although
salt
would
still
be needed.
Molasses-based
supplements
(eg.
molasses/urea
blocks)
will
usually
meet
the
animals'
needs
for
calcium,
sulphur
and trace
elements.
9.4.3
Fermentable
nitrogen supplements
The
main
effect
of a
marked
deficiency
of fermentable
N is
to reduce
feed
intake
(see
Camplinget
al.
1962),
due
to a reduced
rate
of
fermentation
in
the
rumen.
Supplying
urea
to maintain
high
levels
of ammonia
in the
rumen
(ie.
>
200 mg
ammonia
N/litre)
should
optimiseintake,
increase
the
rate
and
extent
of digestion
and
increase
microbial
protein
yield
relative
to
VFA production
(see
Chapters
3 and
4).
Urea
has been
widely
used
as a source
of fermentable
N to
correct
ammonia
deficiency
in the
rumen,
and
although
there
is,
in theory,
a considerable
need
for additional
fermentable
N under
extensive
grazing
conditions,
responses
to urea
supplementation
have
been
unpredictable
(see
Lenget
al. 1973;
Loosli
and McDonald
1968).
In recent
times,
lupins(which are
30%
protein)
have
been
used
successfully
in Australia
to provide
both
fermentable
N and
high
quality
fibreto
grazing
sheep.
One
of the
major
problems
associated
with
supplementing
grazing
cattle
and
sheep
with
urea/molasses
mixtures
is that
nitrogen
deficiency
in the
rumen
is
neither
easily
recognisednor
predicted.
The
N content
of the
pasture
or
faeces,
or even
the moisture
content
of
the faeces,
may
be
indicative
of when
to begin
supplementation.
Ammonia
concentration
in
rumen
fluid is
probably
the
best
guide
(Stephenson
et
al.1984).
A
urea/molasses
mixture
can be
used
to
provide
fermentable
N,
but
there
are
large
differences
among
animals
in the
amount
that
they
consume.
For
in-
stance,
in one
trial,
20
to
50%
of the
sheep
did not
consume
a liquid
urea/molasses
mixture
fed
under
extensive
grazing
conditions
(Nolan
et
al.
1975);
and
20%
of
cattle
did
not
consume
a urea/molasses
block
(R A Leng,
unpublished
data).
In addition,
animals
often
consume
the
mixture
only
sporadically,
which
may
lead
to
large
fluctuations
in rumen
ammonia
levels,
from
above
to
less
than
the optimum
for efficient
rumen
fermentation
(see Chapters
3 and
5). On
the
other
hand lupinsappear
to be
readily
sought
after
by grazing
livestock.
Where
urea
has
been
sprayed
onto
low
nitrogen
pasture
or infused
continuously
into
the
rumen
of grazing
animals,
large
increases
in the
intake
of the
basal
diet
have
been
observed.
Supplying
urea
continuously
to animals
on
diets
based
on low
quality
pasture
has
increased
intake
of
the basal
feed
but has
seldom
led
to
growth
rates
of much
above
maintenance
(Leng
et
al. 1977).
Stephenson
et
al.
(1981)
demonstrated
that
supplementing
ewes
grazing
dry
pasture
in a
tropical
semi-arid
environment
with
urea
in the
drinking
water
increased
lamb
survival,
milk
yield
and lamb
growth
(see Table
9.4
and
Table
9.5).
This
is
somewhat
surprising
since
in
theory,
a peak
in rumen
ammonia
after
drinking
would
have
been
followed
by low
ammonia
concentration
during the
rest
of the
day.
However,
it
may
be explained
if rumen
bacteria
are
able
to
store
a pool
of
nitrogenous
materials
to
draw
on
when
rumen
ammonia
levels
are
low.
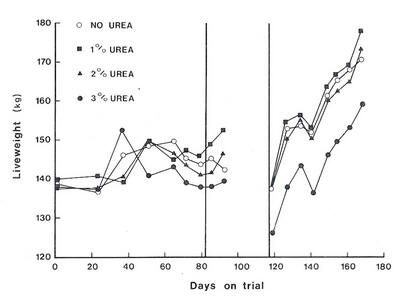
Table
9.4:The
effects
of
supplementation
of
ewes
grazing
dry
native
pasture
with
urea
on
milk
yield,
lamb
mortality
and the
growth
of
surviving
lambs.
No
supplement
Urea
in
drinking
water
(2g/litre)
Ewes
Milk
yield
(ml/4h)
111
123
Lambs
LWt gain
(g/d)
42
76
Mortality
(no.)
9
0
Source:
After
Stephenson
et
al.
(1981).
Table
9.5:
Birth
weight
and liveweight
gain of
lambs,
and feed
and nitrogen
intake,
milk'
yield
and liveweight
loss of
ewes
grazing
low protein, dry Flinders
grass
(hay)
alone (A) or with access to drinking
water
with
2.2g/1
of urea
(B) or fed
Flinders
grass
plus
urea
( C).
Attribute
Hay
(A)
Hay + urea
(B)
Hay + urea
(C)
Ewes
No.
lambing
20
20
20
Feed
intake
(g/d)
900
1190
1250
N intake
(g/d)
8
15
18
LWt change
(kg)
-12
- 8
- 9
Ewes
milked
11
10
Milk yield
(ml/4h)#
60
ND
94
Lambs
Mortality
(%)
40
20
20
Birth weight (kg)
2.9
3.2
3.2
LW
gain (g/d)
35
81
84
Source:
After
Stephenson
et
al.
(1981)
#Mean
yields
measured
on
days 1,
11
and
21
With
ewes
fed
dry,
low
protein
pasture
hay
the
birth
weight
of
the lambs
was
increased
from
2.9 to
3.2kg
by
supplementation
with
urea
either
in
the
feed
or in
the
drinking
water
(Stephenson
et
al. 1981)
(Table
9.5).
In Northern
Australia
recommendations
to
put
urea
in drinking
water
are now
made
on
the basis
of the
rumen
ammonia
eoncentration
in a sample
of
6 to
10
sheep
from
a grazing
flock.
The
value
of this
approach
is illustrated
in Table
9.6.
Table
9.6:
Rumen
fluid
ammonia
concentration
and
milk
yield
of
unsupplemented
and
urea-supplemented
ewes
(urea
in drinking
water).
Rumen
fluid
ammonia
mg/100ml
Milk yield
of
ewe
Lactating
ewe
+ urea
5.1
540
No urea
1.9
440
Non-lactating
ewe
+ urea
6.6
No urea
5.7
Source: Stephenson et al 1984
Stephenson
and
Bird
(1987)
have
recently
taken
their
research
further.
\\'ith
pregnant
ewes
under
extensive
grazing
of dry
native
pastures
and force-fed
supplements
daily
from
115
days
pregnant,
the
researchers
showed
that
urea/ammonium
sulphate
supplements
were
effective
in increasing
lamb
birth
weight
but
found
that
addition
of
meat
meal
with
molasses
further
increased
lamb
birth
weight
(Table
9.7).
Table
9.7:
The
effects
on grazing
ewes
of
N supplementation.
Each
ewe
was drenched
daily
from
115
days
pregnant
with
one
of
the
following:
water,
a
solution
of
urea
plus
ammonium
sulphate(7
+
4)g
(UI5)
or
a
mixture
of
urea
+
meat
meal
+
molasses
(3
+ 100
+
100)9
(lJ15
+
MM).
Supplements
None
U/S
U/S+MM
Ewe
LW
change
(g/d)
38
80
131
Rumen
NH3(mM/litre)
7
10
9
Lamb birth Wt.
(kg)
3.5
3.7
4.0
Source:
Stephenson
and Bird
(1987).
The
difficulties
of ensuring
regular
intake
of
urea
by ruminants
at
pasture
suggest
that
at times
it
might
be advantageous
to
use
protein
meals,
or even
high
protein
grains
such
as
lupins,
to provide
additional
fermentable
N.
The
fermentable
N can
be supplied
directly
by
feeding
a source
of protein
that is slowly
but steadily
fermented.
At
the same
time,
the
amount
of urea
entering
the
rumen
indirectly
from
blood
is increased
(the
extra
urea
arises
from
the
deamination
of
absorbed
amino
acids).
Under
these
conditions
protein
meals
have
many
roles,
ie.
to supply
dietary
amino
acids
for
absorption
and to provide
sources
of slowly
available
fermentable
N,
amino
acids
and peptides
for
microbial
growth
in the
rumen.
9.4.4 Bypass nutrients supplements
Although
the
needs
for
fermentable
N and
bypass
protein
have
only
been
quantitatively
determined
in recent
years,
it has
long
been
recognised
that
feeding
a protein
meal
to
grazing
animals
in the
dry
season
is highly
beneficial.
Marston
(1932)
demonstrated
that
supplementation
with
blood
meal
markedly
increased
wool
growth
of
sheep
on
dry pastures
in
northern
Australia,
and Murray
and Romyn
and their
co-workers
in Zimbabwe
(Murray
€i
al.
1936;
Murray
and
Romyn
1937,
1939)
showed
that the
annual
decline
in condition
of
range
cattle
during
the
dry
winter
months
was
due
to a deficiency
of protein
in the
pasture
during
the
earlier
winter
months
and
to a deficiency
of both
protein
and
energy
during
the
later
part of
the winter.
However,
the
latter
conclusion
may perhaps
be
modified
as they
failed
to provide
fermentable
N,
which
modifies
the
effects
of the
protein
meal.
Figure
9.1
shows
a typical
effect
of supplementing
cattle
on dry
winter feed
with bypass
protein.
Hennessy
et
al. (1981) and Lindsay
and Loxton
(1981)
have
since
shown
that
giving
a bypass
protein
meal
to cattle
on
a diet of
dry Carpet
grass
hay
and
dry
Spear
grass
hay
(45%
digestible,
about
0.5%
N) substantially
increased
the
feed
intake
of the
cattle.
The
results
of
two experiments
in which
urea/sulphur
and
bypass
proteins
were given
to
growing
Hereford
heifers
and pregnant
Brahman cross
cows,
respectively,
are
shown
in
Table
9.8
and Table
9.9.
Table 9.8:
Dry
matter
intake
and liveweight
change
of cattle
(170kg liveweight)
fed
speargrass
hay (predominantly
Heteropogon
contortus) supplemented
with urea
and/01'
f01'1naldehyde-protected
cottonseed
meal (protected-C5M)
Dry matter
intake (kg/d)
LWt change
(kg/d)
Hay
+ urea
3.01
-0.32
Hay
+
protected CSM
3.72
+0.11
Hay
+
protected CSM + urea
4.43
+0.22
Source:
Lindsay et
al.
(1981)
Table
9.9:
Mean
liveweight
change
and dry
matter
intake
of pregnant
cows
(415kg liveweight)
fed
spea1'grass
(hay)
supplemented
with urea/sulphur
(U /5)
and a
bypass
protein
meal
(P P).
The
experiment
was carried
out
over
the last 60d of pregnancy;
the hay was
of low digestibility
and contained
0.4
% N.
Hay DM
LWt
Calf birth
intake
change
weight
Diet
(kg/d)
(kg/d)
(kg)
Spear
grass
4.2
-0.8]
22
Spear
grass+U-S
6.2
-0.31
31
Spear
grass+U-S+PP
8.1
+0.75
32
U-S supplied
55g
Njd.
PP supplied 1kgjd
of a protein
pellet
containing
80
% cottonseed
meal,
10%
fish meal
and
10% meat
meal
(protected
with formaldehyde)
Under
subtropical
grazing
conditions,
similar
responses
have
been
observed
in cattle
given
supplements
of bypass
protein
in the
absence
of
a fermentable
N supplement
(Table
9.10).
Due
to their
greater
weight
gain,
heifers
that
received
supplements
during
the
dry
season
reached
sexual
maturity
one
year
earlier
than
the unsupplemented
heifers.
The
age
of heifers
at first
calving
and
their
fertility
in subsequent
years
obviously
have
a large
effect
on
the animals'
productivity.
Table
9.10: The
effects
on the
mature
body
size
of
protein
supplementation
(during
dry season)
of cattle
. All
calves
were
the
same
weight
in
1977.
Mean live weight, kg
1978
1981
1982
Feeding
system:
Native
pasture
(no
supplement)
197
329
320
Native
pasture
plus bypass
protein
(during dry
season)
Group
1
259
378
382
Group 2#
264
397
397
Source:
Hennessy
(1984).
Improvement
of lifetime
productivity
of
cows
could
be the
most
important
determinant
of
whether
supplements
should
be
fed
to cows
grazing
dry
tropical
or
subtropical
pastures.
Supplementation
with protein
concentrates
in the
dry
season
significantly
increased
the
mature
bodyweight
of the
cows
(Table
9.10),
which
is of
considerable
importance
where
the
objective
of the
system
is to
produce
work
oxen.
It appears
to
be
likely
that
under
many conditions
draught
oxen
are
stunted
by
protein
malnutrition
in early
life.
Research
needs
to be
conducted
into
whether
periods
of
protein
deficiency
in early
life
also
reduce
reproductive
capacity
and/or
milk production.
During
the
pasture
growing
season,
feed
intake of
lactating
animals
on tropical pasture
may
be limited
by the
availability
of
dietary
bypass
protein
from
the pasture.
Stobbs
et
al. (1977)
showed
that
supplementing
cows
on a su btropical
pasture
(largely
Rhodes
grass)
with 1kg/d
of
formaldehyde-treated
casein
(ie.
protected
from
rumen
degradation)
increased
milk yield
(Table
9.11).
The effect
was
attributed
to increased
pasture
intake.
These
results
indicate
that
the
amount
of
essential
amino
acids
absorbed
from
the
digestive
tract
of
animals
on
such
pastures
is
less
than
that
needed
to
support
high
levels
of milk production.
Table 9.11:
Effects of
supplementation
with l kg/d
protected
(formaldehyde)
or unprotected
casein
on yields
of cows
grazing
Rhodes
grass pasture
which
contained
3%
N.
Milk
Supplement
Yield,
kg/d
Fat, %
Protein,
%
None
12.3
5.1
3.3
+ 1 kg casein
12.7
5.2
3.3
+ 1 kg
protected casein
14.7
4.8
3.5
Source:
Stobbs et al 1977
9.4.5 Other supplements
The
possibility
of
using
supplements
with a low
'rumen
load'
and which contain
high
levels
of digestible
carbohydrates
or protein
appears
to
be
a new
concept.
Such
supplements
would
leave
the
rumen
quickly
and have
little
effect
on
rumen
distension
and
therefore
intake
of the
basal
feed
should
remain
unchanged.
Inaddition,
energy
and
protein
that
avoids
fermentation
and is
digested
in the
lower
intestines
is
used
more
efficiently
by
the
animal
than
when
digested
in the
rumen.
Studies
by
Elliott
et
al. (1978a)
showed
that,
in cattle
on sugarcane-based
diets,
rice
grains
in a
rice
polishings
byproduct
left
the rumen
quickly
and intact,
resulting
in increased
total
feed
intake.
Glucose
availability
was
markedly
increased
by supplementation
with this
feed
(Ferreiro
et
al. 1979).
There
is
an urgent
need
to develop
methods
of feed
processing
that
guarantee
rapid
passage
of
such
feeds
through
the rumen.
Feeds
processed
in such
a way
would
supply
essential
nutrients
to
balance
those
available
largely
from
rumen
fermentation.
Such
supplements
are
unlikely
to
reduce
the
digestibility
of cellulose
in the
rumen.
Lupins and cottonseed meal
Inthe
last
five
years
considerable
interest
has
arisen
because
of the
large
quantities
of lupins
that have
been
successfully
grown
and harvested
in Australia.
They
are
finding
an increasing
role
in supplementary
feeding
of cattle
and sheep
when
pastures
are dry
and low
in protein
and digestibility.
In general,
it seems
that lupins fed
at levels
compatible
with their use
as
a supplement
are providing only a fermentable
N source
but because
they are
low in
starch,
easily
handled and easily
fed they have advantages for
feeding
under
extensive
grazing conditions.
The results
of two selected
feeding
trials are
shown in Table 9.12
which clearly
indicates
the different
roles of these
two supplements.
A c.ombination
of
cottonseed
meal
and lupins
is a highly
efficient
way of
providing both fermentable
N and
bypass
protein.
Table
9.12: Lupins (Lup) and cottonseed
meal
(CSM)
are excellent
N supplements
for ruminants on dry
pasture
(5-7
% crude
protein and
43- 49 % digestibility).
The
effects
of lupins
are most likely
to
be through
providing
fermentable
N and
carbohydrates
whereas
cottonseed meal
appears
to be
largely
a source
of bypass
protein.
Species/Supplement
Supplement
intake
(g/d)
% increase
in
pasture
intake
LW change
(g/day)
Sheep
Nil
-
-
-76
Lup
85
42
1
CSM
75
67
61
Cattle
Nil
-
-
-214
Lup
900
11
119
CSM
900
41
321
Smith and Kenney 1987
9.4.6
Dietary history, physiological
state and supplementary feeding on dry pastures
Although
there
is
little
information
on the
ability of cattle
to
utilise
body
protein
reserves,
there
is circumstantial
evidence
that
suggests
that the
"protein
status" of
an animal may
affect
its
response
to energy
or bypass
protein
supplements
at pasture.
Andrews
and 0rskov (1970
a, b) showed
that the
protein content
of the
body
can
be altered
by the level
of bypass protein in the
diet of lambs
grown to a standard liveweight,
and the
ratio of protein to
energy
in the carcass varied
according to the diet. There
is evidence
that suggests
that lambs could grow
at
a high rate
when given
only
a moderate
level of bypass protein
(4%)
in their
diets after
they
had been
kept
at a constant
bodyweight
for some
time (R A Leng,
unpublished
observations).
Body condition
as affected
by dietary
history
of animals
should
be taken
into
account
when
assessing
the effects
of bypass
protein
supplements.
Short
term
studies can
be misleading,
particularly
if the
animals
.n unknown
dietary
history
or they
have
previously been
fed high protein
diets. The apparent
over effects
of feeding
bypass
protein
to cattle
on molasses-based
diets
observed
by Smith
et
al
(1979) may
have
been due
to a lower
requirement
for
protein
relative
to energy
following
a period
of
high
protein nutrition.
A further consideration
is that
an increased
demand for
amino
acids,
as
occurs
in late
pregnancy
and early
lactation,
might
also
modify
a response
to mentation
with
bypass protein.
Both
pregnant
ctating
cattle
increased
their
intake
of a basal
'otein
diet
when
given
a supplemental
bypass
1 (Table
9.13),
but only
by as
much as
young growing cattle.
In
Hennessy's
studies
(Table
9.10),
cattle
had a
higher
intake
of the
basal diet
prior
to supplementation.
Table 9.13: Effect of supplementation (Suppl) with various
protected or bypass protein concentrates (P P) on percentage
intake of native pasture (% increase) by cattle.
Supplement
Hay intake (g/kg0.75/d
% increase in hay intake*
Growing#
Nil
69
PP
99
43
Lactating##
Nil
52
U-S
64
25
U-S + PP
94
96
Pregnant###
Nil
47
U-S
69
47
U-S + PP
91
94
Source: #Hennessy et al 1981; ##Loxton (1981). ### Lindsay et
al. (1982).
U-S: Urea/sulphur supplying 50g N/d.; P P: 1200g of bypass
protein pellet/d.
+
Expressed as a percentage of hay intake of unsupplemented
9.5 TRADITIONAL
SYSTEMS BASED ON PASTURE
9.5.1 Communal
grazing
In many
countries,
traditional
communal
grazing
allows the
animals to
graze
common
land. They
are
housed at
night,
either
from
fear
of predation
by wild animals, of from
fear of theft
or
because
of the
need
to supervise (shepherd)
the
animals.
This
system
is usually
associated
with
subsistence
farming
and livestock
play a
complementary
role
to crop
production,
in
general,
using
the
crop
residues
and
grazing
lands
that
cannot
be cultivated.
The latter
comprise
rocky
hillsides,
highly
eroded
gullies,
roadsides,
areas
around
cultivated
plots,
the
bottoms
of valleys
that
are too wet
to be
cultivated
and areas
set aside
for
communal
use. This
pattern
of
livestock
production
is most
common
in
areas
with
relatively
high
rainfall
and
often
marked alternating
wet and
dry
seasons.
These
conditions
are typical of the
heavily
populated
and
intensively
cropped
humid
tropics.
Usually
a variety
of animals
are
involved
in a mixed
grazing
group
which
is invariably
shepherded.
Use is
sometimes
made of
communal
lands
by tethered
animals
which
are
moved
according
to the
availability
of pasture.
The
animals
are
usually
grazed
for 8 to 10 hours, from
early
morning
until
late afternoon.
The
pasture
is
usually
sparse
and,
because
of the
pressure
from large
numbers
of animals,
the
availability
of feed
biomass
is
often
low.
For these
reasons
animals
are
generally
fed
cereal
straws
in the
corral at
night
and obtain
an
appreciable
proportion
of their
diet
from
this
source.
Overgrazing
has been
considered
to be
a negative
attribute
of
these
systems. However, overgrazed
pasture
provides
young,
highly
digestible
herbage
often
high in fermentable
N.
The
sharing
of this
pasture
by large
numbers
of animals
provides
the high
quality supplement
required
to increase
the
efficiency
of utilisation
of the
cereal
straws
(Chapter
8)
which
often
form
the largest
part of
the animals'
diet.
Since
the
animals'
intake
of the
green
pasture
is
likely
to
be
low
and not
synchronised
with the
ingestion
of
crop
residues,
it is
likely
that
additional
supplements
providing
fermentable
N (eg.
urea)
or
bypass
nutrients
or both
could
increase
productivity.
In the
dry
season
the availability
of pasture
on communal
land
is much
lower
than
in the
wet
season
and
the
fermentable
N intake
of grazing
animals
will be
inadequate.
In these
circumstances
small supplements
oflegume
hay
to
provide
bypass
protein
and fermentable
cellulose
may
be
of considerable
benefit
and responses
to urea
and bypass
protein
are
to
be
expected.
During the
dry
season,
grazing
animals
will benefit
from
continuous
access
to
fermentable
N supplements
and this
should be
the
basis
of any
strategy
to
improve
individual
animal
productivity.
9.5.2 Cut-and-carry grass
Specialised
systems
have
developed in which forage
crops
are grown,
harvested
and carried
to the
animals.
These
systems
are generally
used only
where
high prices
are
paid for
animal products,
usually milk, or where
the
stock
are valuable
as a basis of breeding.
These
systems
are
often
justified
on the
basis
of the
very
high
yields
of dry matter
that are
obtained
when
tall growing forages
(eg.
elephant
grass) are
heavily
fertilized
(and
frequently
irrigated)
and harvested
at frequent
intervals.
In practice,
the forage is
usually
harvested
at an advanced
stage of
maturity
and is highly
lignified
and of
low digestibility;
its feeding
value
is thus
little
better
(perhaps
not
as good)
than that
of cereal
straw.
Smallholders
in some
countries
supplement
their
crop
residues
with mixed
forages
and tree
foliage
gathered
from
roadsides,
dykes,
bunds
and areas
not grazed
by
livestock.
In some
specialised
systems
a 'proportion'
(5%)
of the
cropping
land
is set
aside
to
specifically
produce
high
protein
legumes
to
supplement
crop
residues
(eg.
the
Loess
Plateau
of
Northern
China).
These
forages
are
given
in small quantities
and serve
a useful
function in providing
some
of the
nutrients
that may
be
otherwise
deficient
in the
basal
diet.
They
rarely,
however,
provide
sufficient
fermentable
N and
little
bypass
protein
for the
utilisation
of
a mainly straw-based
diet.
These
systems
relate
to
most
smallholder
family farms in developing
countries
and therefore
should
be a target
for
the introduction
of
innovations
such
as:
Using solidified
multi-nutrient
blocks
(Chapter
11) and finding simple
methods
for
protecting
the
protein
in the forage
supplements
Upgrading
the
forage
supplement
by
establishing
legume
trees.
Examples
of this
are the
planting
of ipil-ipil (leucaena)
on roadsides
in the
Philippines
and the planting
of
communal
land with tree
crops
in India.
The
advantage
of tree
crops
over
grasses
and low
growing
legumes
is
the
multi purpose
nature
of the
former,
which
provide
forage,
fuel
and
timber
Introducing
a proportion
of
tanniferous
legumes
or trees
(2-4%
tannins)
into the
forage
supplements.
9.5.3 Transhumance systems
Large areas
of Africa
and Asia
have
been used
for
grazing
livestock
since
these
were
domesticated
about
10,000
years
ago.
The
grazing
areas
around
villages and
along
established
nomadic
routes
are
communal
and
a mixture
of pasture
plants
have
become
adapted
to these
grazing
systems
(Chalmers
1976).
Although
pastoralism
is diminishing
in some
regions
because
of political
pressure
or expansion
of
cropping
areas
or a combination
of the
two,
it is
still
the
only
feasible
means
of exploiting
plant
growth
on
relatively
poor land
under
harsh,
usually
arid,
environments.
The system
has
survived
for
a long
time
and
supports
a large
population
of
animals
on which
a large
number
of
people
depend.
Nomadic
or transhumance
grazing
systems
exploit
the
forage
resources
of different
rangelands
when
each
is at its best.
Often
the
rangelands
are
grazed
until
they
are
cleared
of accessible
biomass.
More
often,
the
rangelands
are abandoned
for
greener
pasture
when
some
biomass
of
fermentable
carbohydrate
is still
available.
This
biomass
is
used
inefficiently
because
of the deficiency
of
essential
nutrients
(ie.
fermentable
N),
and
the
animals
lose
weight.
Pastoralist
systems
are
the
most difficult
in which
to apply
scientific
feeding
principles.
Despite
this,
animal
productivity
per
hectare
from
these
traditional
systems
is often
greater
than
that
from
ranching,
especially
when
account
is
taken
of
the contribution
of
meat,
milk
and
blood
to the
human
diet.
There
is
great
scope
for
extending
the existing
trends
of integrating
transhumance
systems
with subsistence
cropping,
since
the
pastoralist
benefits
from
the
crop
residues
in the
dry
season (when
grazing
is
most deficient)
and
the
crop
farmer
uses
the
animal
excreta
as fertiliser.
This
system may
be further
improved
by
the
application
of principles
discussed
in
this
chapter.
For
example
if the
crop
farmers
could
be persuaded
to grow
a legume
bank (particularly
one
containing
tanniniferous
legumes)
in a crop
rotation,
he
would
benefit
in two ways:
first,
the
legume
would provide
nitrogen
for a subsequent
crop
and second,
the
legume
would
be a cash
crop
for
'sale'
to the
pastoralist.
The
pastoralist
benefits
from
the catalytic
effect
of the
legume
forage
in improving
the
utilisation
of
the
crop
residues
by
his
livestock
.
In recent
years
pressure
from
an increasing
population
has led
to overstocking,
which
tends
to
"mine"
the
fertility
of the
soil
and
no efforts
have
been
made to maintain
or
increase
the
fertility
of these
pastures
(a necessity
for
long
term
stability).
The
solution
to these
problems
is
to improve
traditional
transhumance
systems
and not
to discard
them
in favour of
"exotic"
systems.
In
these
grazing
systems
the
objective
must
be to
increase
the
efficiency
of feed
conversion
to products
rather
than
to
increase
the
intake
of
pasture.
Supplementation
with
urea and
minerals
is
the
only
strategy
that should be
recommended
unless
stocking
rates
are
reduced,
or
pasture
biomass
is
increased
by fertiliser
application.
Under
these
conditions
additional
supplements
can
be
used
to boost
intake
and
feed
conversion
efficiency.
9.8 Drought feeding
9.6.1 Introduction
In many
countries
farmers
have
to
contend
with
periodic
droughts.
The
transhumance
migration
patterns
were
largely
a result
of
annual
shortages
offeed,
due
to either
drought
or temperatures
being
too low
for plant
growth.
In large
continents
where
seasons,
and
therefore
the
availability
of pasture
(or
grazing),
tended
to 'move'
across
the
continent,
wildlife
moved
with
the
availability
of green
pastures
and their
reproductive
patterns
were
synchronised
to maximise
the
benefits
from
the
best
pastures.
The
migrations
of wildlife
in Africa
and
of
the Caribou
(to
the
Arctic
in summer
and back
to the
winter
grazing
at the lower
latitudes)
are
examples
of this.
Information
is
now
available
that
permits
rational
supplementation
of
dry pastures
for more
efficient
production.
However,
in annual
or short term
droughts
particularly
where
animals
are prevented
from
migrating,
management
practices
have
to be developed
in which
some
of
the
pasture
is un grazed
during
the
growing
season
and
retained
for
use
in
drought.
Suitable
supplements
must
be
made
available
to maximise
the
efficiency
with
which
this stored
pasture
is
utilised.
Conserved
pasture
should
contain
legumes
if possible,
as this
will reduce
but not
eliminate
the
need
for
supplements.
Where
livestock
are
integrated
with crop
production,
cereal
crop
residues
and byproducts
can be
stored
for
drought
feeding.
9.6.2 Water economy
There
is an
extensive
literature
on the
water
economy
of herbivoro'us
livestock
(Macfarlane
1964;
Louw
1984).
It is
not
the
objective
here
to
discuss
all the
factors
that influence
water
utilisation
by ruminants,
but rather
to highlight
those
features
which
relate
specifically
to
the
effects
on efficiency
of digestion
and metabolism.
Water
is
essential
for
ruminant
performance.
It
is needed
for
milk
production
and
for
excretion
of
salts
in
urine
and
faeces.
Requirements
for
water
are
related
to
heat
load
and
level
of
production.
Water
intake
appears
to be
well
controlled
and closely
corresponds.
to requirements.
Animals
grazing
succulent,
young
pasture
may
not
drink,
whereas
water
intake
is
optimised
when
pastures
are dry
or where
feeds
contain
little
free
water.
Water
is
required
in growth
and total
body
water
tends
to be
related
to body
protein
content;
it is
obviously
required
for
milk production
(Louw
1984).
Water
cannot
be regarded
as a nutrient
but
there
is
a high
correlation
between
water
and
dry matter
intake,
sheep
consume
between
1.5
and 3 litres
of water
per
kg of
feed
dry matter
in the
absence
of
a heat
stress
or a salt
load.
Comparable
requirements
for
cattle'
are
2.5
to
4.5
litres
per
kg
of
dry
matter
intake
(see
Phillips
1960).
When
water
is restricted
the ratio
may
fall
to as
low
as 1.5
litres
of
water
intake
per
kg of
feed.
Conversely,
the
ratio
tends
to rise
if
feed
intake
is
restricted.
When
nutrients
are imbalanced
and there
is
a "burn
off"
of
excess
acetogenic
substrate,
the
imposed
heat
load on
the
animal
may become
a constraint
where
animals
are
stressed
by high
ambient
temperatures/humidity,
and
this
stress
will be
increased
if water
is restricted.
The
problem
of
water
intake
and
dissipation
of
metabolic
heat
is compounded
in lactating
animals,
because
of their
higher
feed
intake
and
high
metabolic
rate.
All animals appear
to be able
to reduce
urine
volume
and
to excrete
concentrated
urine
when
water
is restricted.
There
appears
to be
considerable
genetic
variability
in the
capacity
to concentrate
urine
both
within
breeds
and between
breeds
(see
Phillips
1960).
Effects of restriction on deprivation of
water
Restriction
of
water
has immediate
and drastic
effects,
setting
in train
the
following
series
of events.
Reduqtion
of
urine
volume
until the maximum
capacity
to
concentrate
the
urine
is reached
Reduction
in feed
intake-which
also reduces
the
urine
volume
and
obligatory
water
loss
in faeces
Reduction
in total
body
water-primarily
extracellular
water-and
haemoconcentration,
along
with
some
build up
of
sodium
and
chloride
ions
If water
deprivation
is severe
then
dehydration
occurs,
there
is
an increase
in
protein
catabolism
and finally,
failure
of renal
function.
In
arid
areas,
tolerance
to
dehydration
is an
extremely
important
physiological
trait,
particularly
where
the
available
feed
is dry
and "imbalanced"
for
nutrients,
and there
is heat
stress.
The
potential
to dehydrate
is also
increased
when
the
animal
has to
work
and
also
to cover
long
distances
between
watering
sites
and feed.
Where
livestock
are
not herded,
the
capacity
to tolerate
heat
stress
and
dehydration
and,
at the
same time
to maintain
feed
intake,
will
determine
production
(or
survival).
Where
animals
are
supervised,
as occurs
in most
developing
countries,
herders
establish
a set
pattern
of
intermittent
watering,
often
daily,
once
every
two
days
or
once
every
three
or four
days.
The
pastoralists
are
usually
skilled
at ensuring
that
the animals
obtain
at least
a minimum
of green
pasture
from
selected
sites
in the
extensive
grazing
areas.
This
has
beneficial
effects
on the
utilisation
of
the
dry
herbage
through
provision
of
some
fermentable
N in
the
rumen
and
a possible
higher
retention
and
recycling
of urea
N to
the
rumen.
Tolerance
of species
and breeds
to water
restriction
Species
vary
widely
in
tolerance
to
dehydration
and
most
animals
die if weight
loss
exceeds
20%
of
body weight.
Exceptions
are
the
camel
and
Merino
sheep
which
appear
to
tolerate
up
to 30%
weight
loss.
The
point
at which
feed
intake
begins
to be
restricted
by
water
deprivation
is an
all important
characteristic
which
may
determine
an
animal's capacity
to
survive
or produce
under
harsh
grazing
conditions.
It
seems
logical
that
animals
with
a relatively
low
basal metabolic
rate
(and
therefore
lower
heat
production)
will have
an advantage
in the
dry
topics.
Obviously,
the
less
water
that
is
needed
for
thermo
regulation
the more
will be
available
for
osmoregulation.
Zebu
cattle
use
water
more
efficiently
than
European
breeds
(Fourie
et
al1968)
perhaps
because
of their
lower
basal metabolic
rate
(Vercoe
and Frisch
1982).
The
capacity
of some
ruminant species
to produce
high
fat milk (an acetogenic
sink which
reduces
the
animal's
heat
production)
also
confers
metabolic
advantages
in situations
where
water
restriction
is associated
with
predominantly
dry
grazing
(giving
rise
to
imbalanced
nutrients).
In
this
respect
the buffalo,
indigenous
cattle
breeds
(Zebu,
Ndama,
Criollo),
camel
and Merino
sheep
have
obvious
advantages.
Frequency
of watering
and
nutrient
utilization
Scientists
often
read
much
physiological
significance
into
the
effects
of
intermittent
watering
on ruminant
productivity.
However,
the
practical
advantages
of intermittent
watering
appears
to
be well
understood
by pastoralists
in arid
regions.
These
people
have
developed
watering
regimes
which
optimise
the
conflicting
requirements
for
minimising
livestock
losses
and the
cost
in time
and energy
expenditure
of moving
their stock
over
long
distances,
set against
a background
of
sparse
availability
and low
nutritive
value
of feed.
Extended
watering
frequencies
that
have
proved
to be
most
practical
(usually
once
every
three
to four
days)
have
advantages
of reducing
overall
feed
and
water
consumption
with
improved
nutritional
benefits
in terms
of
increased
feed
digestibility
(see
More
and Sahni
1981).
The
effects
on
digestibility
are
due
primarily
to increased
retention
time
of
digesta
in the
rumen
(Brosh
et
al. 1986a).
Production
traits
(maintaining
body
reserves)
appeared
to be
unaffected
in
animals
watered
every
three
days
as
compared
with
a daily
watering
frequency
(Brosh
et
ai.
1986b).
An important
nutritional
advantage
from
intermittent
watering
may
be the
more
efficient
return
of urea
to
the rumen
improving
the status
ofthe
rumen
microbial
ecosystem.
On
low
N forages,
sheep
are able
to retain
a greater
proportion
of
the
urea
produced
in the
body
(Cocimano
and
Leng
1966)
and when
water
is restricted
a greater
proportion
of
this
urea
may
eventually
reach
the
rumen
for
use
by microorganisms
(see
Ikhantua
et
al.
1985).
Conclusion
Water
is
essential
for ruminant
production.
The
requirements
for
water
are
related
to heat
load,
growth
and for
milk
production,
and
for
excretion
of
salts
in urine
and
faeces.
There
is
a high
correlation
bet
ween
free
choice
intake
of feed
dry
matter
and water.
Generally
ruminants
take
more
water
than
is needed
and adapted
animals
can
often
get their
water
requirements
in a single
drink
given
ei
ther
daily
or once
every
three
or
four
days.
If supplementation
of
dry pasture
is
undertaken,
water
intake
may need
to
be more
often
because
of the
increased
feed
intake.
The
relationship
between
feed
intake
and
water
intake
on
balanced
and unbalanced
diets
needs
considerable
research
as the
effects
of
"balancing"
a diet
by supplementation
could
be offset
by the
negative
effects
that
might
result
from
intermittent
watering.
9.6.3
Nutritional aspects of sub-maintenance
feeding
Drought
feeding
of ruminants
is
often
aimed
at survival
or
maintenance.
When
droughts
are
prolonged,
and feed
resources
become
scarce,
"nonconventional"
resources
are
often
mobilised
for
livestock
feeding.
As
the
objective
of these
feeding
systems
is
to
prevent
excessive
weight
loss,
feeding
levels
are
generally
chosen
which
will maintain
the animals.
The
results
of two
trials
on "survival"
feeding
demonstrate
that
the
efficiency
of
use of
the
available
feed
is a function
of
the balance
of nutrients
available
to the
rumen
and
the host
animal.
Molasses
is
deficient
in fermentable
N for
growth
of
rumen
bacteria,
and
therefore
unless
it is supplied
fermentative
digestion
may be
inefficient.
This
is
borne
out
by the
work
of
Gulbransen
(1985) (see
Table
9.14)
which
showed
that
young
cattle
maintained
their
bodyweight
when
given
about
3.2kgjd
of
molasses
plus
urea,
but lost
weight at the
same
molasses
intake
when
urea
was
absent.
It is
concluded
that
the animals
fed
molasses
without
urea
must
have
increased
their
metabolic
heat
from
part
of
the diet
(see
Leng
1986).
Table
9.14:
The
effect
of
providing
fermentable
N (urea)
on the
efficiency
of
utilisation
of molasses
for
body
weight
maintenance
of,
young
cattle.
Live weight,
kg/d
Molasses, kg/d
No urea
+ urea
2
-0.25
-0.14
3.2
-0.12
0
Adapted from
Gulbransen 1985
Diets based on whole cottonseed
In Australia,
whole
cotton
seed
is occasionally
used
as a drought
feed,
due
to
its
ready
availability
and
low
cost
in the
Northern
part
of
the
country
where
it
is
produced.
The
data in Table
9.15
show
that
mature
cows
needed
3kg/d
of
this feed
in order
to maintain
bodyweight.
However,
when
one
third
of the
seed
was
replaced
with
solvent-extracted
cottonseed
meal
(a known
source
of bypass
protein),
the
animals
gained
580g/d.
Apparently,
a feed
mixture
providing
some
bypass
protein
supports
a better
balance
of
nutrients,
even
for
maintenance,
than
one
which
contains
few
bypass
nutrients,
ie.
is degraded
almost
entirely
in
the
rumen.
Table
9.15:
The
effect
of
feeding
a bypass
protein
(cottonseed
meal
CSM)
with
whole
cottonseed
(WCS)
on
live
weightchange
of
mature
cows
under
drought
feeding
conditions.
Live weight, kg
Initial
Final
LW change, g/d
WCS,
3 kg/d
452
461
48
WCS,
2
kg/d
+
CSM,
l kg/d
471
501
575
Source:
R
J
Thompson
and I W Dixon
(1986)
unpublished
observations
Where
droughts
are
seasonal
or
occur
say
one
year
in two,
consideration
can
be given
to
using:
Carry-over
pasture
which
has been
excluded
from
normal
grazing
Cereal
residues
and
byproduets
that
have
been
retained
for
such
eventualities
Bagasse,
cane
tops and
molasses
from
the sugar
industry
In some
situations
hays
or silages
can be
prepared
during
the
pasture
growing
season.
Deterioration
of
dry feed
resources
with
time
is
a major
problem,
particularly
if the
storage
conditions
are
poor.
Although
straw
and
hay
can
be kept
for
3 to
5 years,
in
general
they
are
short
term
storage
products.
On the
other
hand,
silage,
if properly
made
so
that
it is
highly
acidic,
can
be
stored
almost
indefinitely.
Where
drought
recurs
it is difficult
or uneconomic
to make
silage.
In addition,
almost
always when
pasture
is
available,
silage
or
hay making
is difficult
because
of wet
conditions.
If the
drought
affected
area
is
close
to
irrigated
arable
lands,
utilisation
of
the
waste
products
from
intensive
crop
production
should
be encouraged.
These
target
byproducts
include
sugarcane
tops,
bagasse
and molasses
(Chapter
8).
Cane
tops
must
be ensiled
or dried
and briquetted,
bagasse
needs
to be
dried
and
molasses
must
be
stored
to ensure
long
term
availability.
Often
a large
proportion
of
the
byproducts
of
the sugar
industry
is wasted
because
the
main responsibility
of the
management
is
to
ensure
the
efficiency
of the
milling
operations.
Thus,
when
storage
tanks
are
full,
molasses
may
be
disposed
of
into
pits
or rivers,
or
sprayed
on
roads
to
keep
down
dust.
The
use
of
these
feed
resources
in drought
periods
requires
a rational
approach
which will depend
on the
objectives
of the
animal
production
system
and
their
priorities.
These
may
be:
Production
(for
instance
the
production
of
a calf
is often
essential
to ensure
future
viability
of the
enterprise)
Maintenance (as for work oxen)
Survival
(which
is probably
the
only
option
in prolonged
droughts).
More
often
than
not,
and depending
on
the
drought
feed
resource,
the
supplement
can
be chosen
to ensure
maintenance
(urea)
or
production
(minerals,
urea
and
bypass
protein).
The
benefits
of feeding
urea
in a ration
under
simulated
drought
conditions
are
shown
in Table
9.14.
At the
same
low
level
of feed
intake,
urea
supplementation
decreased
bodyweight
loss
of
cattle.
Urea
supplementation
would
have
improved
the
balance
of
nutrients
available.
This
demonstrates
that
the
nutrients
from
rumen
fermentation
in
cattle
on rations
with
adequate
urea
were
better
balanced
to meet
the
animals'
needs
and enabled
them
to use
the
feed
more
efficiently.
Thus the
need
to create
an efficient
rumen
fermentation
is important
ever..
in animals
fed
at less
than
maintenance.
The
data
in Table
9.15
also
indicate
that where
animals
are
restrictively-fed
a drought
diet
the
efficiency
of feed
utilisation
can
be
improved
by
incorporating
a bypass
protein
in the
diet.
A
strategy
for drought feeding
based on the use of
molasses
When
drought
is unexpected
and famine results,
the
primary
limitation
to alleviation
of
the problem
is
the
cost
of
transporting
feeds.
Sources
of feed
are invariably
localised
and usually
in intensive
agricultural
areas,
which
are often
distant
from
the
concentrations
of livestock.
It
is
not
conceivable
in countries
other
than
those
with
small
populations,
such
as
Australia,
that
grains
could
ever
be used
to
feed
animals
during famines.
In examining
the availability of
high
energy-dense
materials,
only
molasses
can be
considered
as a drought
relief
feed
in the
areas
where
famine
is endemic.
The
constraints
to
the
use
of
molasses
are
transportation
and the
availability
of storage
tanks.
The
availability
of molasses
is
seasonal.
Long
term
planning
should
allow
for
the
molasses
to be
transported
during
the
crushing
season
to suitable
storage
points
from
where
it can
be
distributed
as
required.
It is
envisaged
that molasses
will be inexpensive
during the
crushing
season,
particularly when
the
molasses
tanks
are
full.
Tankers
may
be
employed
in this
system
and
storage
points
set
up
in the
areas
prone
to drought.
Storage
should
be
preferably
in areas
of clay
soils,
where
the
molasses
can
be
stored
in
simple
unlined
but covered
pits
in the
ground.
In other
areas
pits
lined
with plastic
sheets
could
be
readily
installed.
In recent
years,
a drought
feeding
strategy
has
developed
for
parts of
Australia
that are
prone
to
drought
using
molasses
(see Nicol
et
al. 1984).
Urea
is
mixed
in the
molasses
(8%
w/w):
this
is more
urea
than
is needed
for
fermentation
of
the molasses
but
the
high
level
restricts
the
intake
of
molasses
and the
surplus
helps
to supplement
any available
forage.
More
than
one
million
animals
were
fed
this
mixture
in the
drought
in Queensland
in 1983/84.
In India the
drought
feeding
strategy
uses
molasses
which
is hand mixed
with bagasse,
urea
and minerals
and fed
as
a survival
ration
to livestock,
which
are
collected
close
to
the
sugar
mills to
reduce
the
need
to transport
the feed.
Under
extensive
grazing
conditions
sheep
and
cattle
appear
to be
able
to
find
enough
roughage
to aid the
utilisation
of
the molasses
and to
maintain
the efficiency
of the
rumen.
In rec.ent
studies
a different
strategy
has
been
developed
based
on
the use
of low
levels
of urea
(2%)
in molasses.
The low
level
of
urea
prevents
exc.essive
intake
of the
mixture
by restricting
the amount
of
nitrogen
available
for
fermentation
in the
rumen
(Ffoulkes
et
al. 1984).
Supplementing
this
diet
with
a bypass
protein
increases
molasses
intake
and
the productivity
of
the animals,
thus
giving
a number
of
husbandry
options.
For
example,
if the
farmer
wants
to
breed
his animals,
the
cows'
body
weight
and/or
balance
of nutrients
may
have
to
be
improved
to ensure
fertility.
In addition,
when
the
drought
breaks
the
animals'
body
condition
can be
rapidly
increased
to optimise
utilisation
of
the pasture
that becomes
available
(see
Figure
9.5).
Figure
9.5: Liveweight
change
of
cattle
fed
ad libitum a survival ration
of molasses
and wheat
straw (0.5%
liveweight)
with 0, 1, 2,
or 3% urea and a growing
ration
with an additional 500g/d
of a bypass protein
supplement.
The
animals were
on pasture
for
one month
between
the survival
and the
growing periods
(Source:
Ffoulkes
et
al.
(1984).
Molasses
blocks
of
relatively
low
urea
content
could
also
be
used
in drought
as an
energy-dense
feed.
Molasses
blocks
are
consumed
slowly
and
intake
is
restricted
by the
hardness
of
the blocks;
this
could
be
important
in conserving
the
primary
feed
resource
where
intake
cannot
easily
be
controlled.
In
some
countries
farmers
can
handle
liquid supplements
easily.
Liquid
supplements
were
used
successfully
in Ethiopia,
where
donkeys
were
used
to distribute
urns
of a molasses/urea
mixture
within
drought
affected
areas
(Preston
and
Lewis
1984)
(Plate
9.1).
In all molasses
feeding
systems
where
the
intake
of feed
is limited,
improved
efficiency
of
utilisation
of the limited feed could
be expected
if a small
quantity
of a bypass
protein
could
be included
in the
diet.
9.6.5
Responsibility
for drought
feeding
Undoubtedly
it is
the farmer's
responsibility
to provide
feed for
livestock through normal
seasons.
However,
in abnormal
seasons,
when droughts
are prolonged
and people
are at risk
and famine
occurs,
the responsibility
for feeding
livestock
must
rest
with
the
government.
Breeding
stock
and oxen
are essential
for the survival
of
people.
Once a drought is over,
crops have to be planted
and depleted animal
populations
have to be restored.
It is in
the
national
interest
that strategies
are
developed
to ensure
the supply
of feed
for
essential
animals
at these times.
9.6.6 Conclusions on drought feeding
In
most
situations
in which
drought
is predictable,
farmers
must
develop
strategies
for
conserving
feed. This
may
be
as standing
pasture
that has
not
been grazed
during
the wet
season,
as hay or
silage or
as cereal
crop
residues.
If the nutrients
available
from
these
resources
are correctly
balanced
feed
utilisationefficiency
can be high.
Close to the areas
of sugar production,
there is potential
for
farmers to store
molasses
either as liquid feed or
as gelled
molasses blocks
to be
used when drought
is prolonged.
Where
droughts
cannot
be predicted,
particularly
where
famine ensues,
it is
the
responsibility
of government
and international
agencies
to assist
in maintaining
feed
supplies
for
animals,
to
allow
farming communities
to survive.
A long term
drought feeding strategy could
be started
in most
tropical countries
almost immediately
using primitive
means of storing molasses, which
could
be dispensed
in times
offamine and its efficiency
of use could be ensured
by
adding
urea
and minimal
amounts of a bypass
protein meal.
It should
be emphasised
here
that no attempt
is made to use the
recommended
nutrient
requirements
for maintenance,
pregnancy
or lactation in
drought feeding.
In
contrast,
the feeding systems
advocated are
often
ad libitum, the
level
of feed intake
being adjusted
by manipulation
of the diet.
The
following
systems
are advocated:
1. Animals
with access to pasture
Maintenance,
survival
or first
6 months
of pregnancy
feed
molasses
containing
8-10%
urea
ad libitumwith
small
amounts
of
a meal
high
in protein
and fat
(50-100g
for sheep
and
250-500g
for
cattle
(this
will
ensure
a high
feed
utilisationefficiency).
Last
trimester
of pregnancy
feed molasses
with 3% urea
ad
libitum plus a meal high
in fat and
protein (eg.
rice polishings,
maize
germ,
copra meal).
2. Lactating
animals
molasses
with 3% urea
ad
libitumplus
500g to I kg daily
of a protein
meal rich
in lipids (eg.
pressure
extracted
oilseed
meal)
depending
on potential
for milk
production.
3.
Animals
enclosed
and without
access to forage:
Where
animals
are
totally
enclosed,
although
the
above systems can be applied,
it
is
suggested that roughage
should
be fed at 0.5%
of bodyweight.
Liquid molasses
could
be partly
or wholely
replaced by molassesIurea/roughage
blocks.
Plat.e9 .1
Molasses
was
distributed
to
drought areas
by tanker
but
reached
the
small
farmers
carried in
urns on
the
back
of donkeys
(ILCA,
1985)
Molasses-urea blocks being fed to cattle under drought
conditions in North Africa (Sansoucy R)